Research
1. Secondary metabolism/natural products:
a. The role of metabolites produced by nonribosomal peptide synthetases or polyketide synthases in the life of the fungal cell and in both necrotrophic and hemibiotrophic interactions with plant hosts.
Our lab is involved in an exhaustive functional characterization of the set of non-ribosomal peptide synthetase (NRPS) and polyketide synthase (PKS) genes of the corn pathogens, Cochliobolus heterostrophus and Setosphaeria turcica, and the small molecules produced by the enzymes they encode, to ascertain what the metabolites are doing for the fungal cell. To date, the NRPS method of peptide biosynthesis has been described for filamentous fungi and bacteria, only. In addition to vast structural diversity, non-ribosomal peptides (NRPs) and polyketides (PKs) have a broad spectrum of biological activities, many useful in medicine, agriculture, industry, and biological research. However, to suggest that inter-organismal activity is their primary function is likely incorrect; in our lab, we strive to understand the physiological significance of these peptides to the producing fungi themselves. We hypothesize that these molecules play more diverse roles than previously revealed and that, in some cases, the NRPSs and PKSs are purveyors of small molecules for basal metabolism. These molecules are required also in particular environmental niches, often, but not always, associated with stress conditions [e.g., low or high iron, oxidative or nitrosative stress, on the plant host, when mating, when the fungal population is too high (or too low), etc]. Our goal is to understand the mechanisms by which these small molecules impact fungal development and effect environmental responses, using functional analyses.
b. Extracellular and intracellular siderophores; roles in iron homeostasis and fungal development (siderophores, sex, and virulence).
Extracellular and intracellular siderophores; roles in iron homeostasis and fungal development (siderophores, sex, and virulence). Iron is essential for most organisms, however, bio-available forms are limited. Efficient iron-uptake/storage mechanisms are critical to survival. For this, fungi employ a variety of mechanisms, including siderophore-assisted mobilization, the focus of this project. Siderophores, high-affinity iron chelators, function in iron acquisition and sequestration and are biosynthesized by nonribosomal peptide synthetases (see Project 1). Intracellular siderophores are antioxidants, preventing the Fenton reaction, which generates reactive oxygen species (ROS), in the presence of free iron. To date, siderophore biosynthesis has been described for fungi, bacteria, plants, but not for animals. Siderophore-mediated virulence has been demonstrated for opportunistic human pathogens and for filamentous ascomycete phytopathogens. Mutant phytopathogenic fungal strains, lacking extracellular siderophores, are also increased in sensitivity to iron depletion and to ROS, coupling iron, oxidative stress, and virulence. Our laboratory has demonstrated that intracellular siderophores play a seminal role in sexual reproduction. Building on our discoveries, we are dissecting siderophore biosynthetic pathways in Cochliobolus heterostrophus, Setosphaeria turcica, and in Fusarium graminearum/Gibberella zeae, to capture genes whose expression depends on integrity of the siderophore pathway. We propose iron as a key player in plant-fungal interactions; iron availability impacts ability to cause disease and to disseminate in the field.

Fusarium graminearum strains lacking both intra and extracellular siderophores (Right) cause almost no disease.
c. Unraveling the genetic mechanism responsible for production of T-toxin, a host-selective polyketide toxin, required for high virulence of C. heterostrophus race T to corn.
Cochliobolus heterostrophus, ourmain experimental organism, is an especially appropriate subject for a study of genes required for biosynthesis of nonribosomal synthetases or polyketide synthases, because it belongs to a genus renowned for its ability to produce secondary metabolites. Closely related genera include the Alternaria, Setosphaeria, and Pyrenophora spp., genera best known for their host selective-toxin producing abilities. Among these, C. heterostrophus is unsurpassed in terms of genetic tractability and very efficient homologous recombination between transforming DNA and target genomic sequences, which facilitates functional analysis by site-specific gene deletion. The genomes of several strains of C. heterostrophus race T and race O, as well as other Cochliobolus species have been sequenced by the Joint Genome Institute.
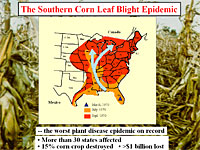
Fungal pathogenesis to plants is an intricate developmental process requiring biological components found in most fungi, as well as factors that are unique to a particular fungal-plant interaction. The Tox1 locus, which controls biosynthesis of the host-selective toxin known as T-toxin in C. heterostrophus race T, a pathogen of maize, is an intriguing example of the latter type of virulence determinant. Originally defined as a single genetic locus, it is, in fact, two exceedingly complex loci on two chromosomes that are reciprocally translocated with respect to their counterparts in weakly pathogenic race O which does not produce T-toxin. Highly virulent race T was first identified when it caused an epidemic of Southern Corn Leaf Blight (SCLB), which devastated the US corn crop in 1970. Texas male sterile cytoplasm (T-cytoplasm/Tcms) corn, widely planted at that time, is exquisitely sensitive to race T.
The SCLB epidemic of 1970 generated enormous concern not just about the short term effects on growers’ incomes and corn market prices, but also about the overall “genetic vulnerability” of our essential crop plants. Excerpts from an article by J. Doyle (Doyle, 1985) serve to illustrate the SCLB impact:
… “ In the summer of 1968, … the first signs of trouble went almost unnoticed. Out in the heartland, on a few isolated seed farms in Illinois and Iowa, a mysterious disease was producing "ear rot" on corn plants. “…
…“Reproducing rapidly in the unusually warm and moist weather of 1970, its spores carried on the wind, the new disease began moving northward toward a full-scale invasion of America's vast corn empire. Later to be identified as "race T" of the fungus Helminthosporium maydis, it soon became known as the Southern Corn Leaf Blight.”…
…“The new fungus moved like wildfire through one corn field after another. In some cases it would wipe out an entire stand of corn in ten days. …. In extreme infections, whole ears of corn would fall to the ground and crumble at the touch.”…
…“Nevertheless, with the opening of business that Monday, panic gripped the commodity and futures markets. Estimates that the blight might wipe out half the nation's corn crop fueled frantic trading and speculation. At the Chicago Board of Trade, the nation's largest commodities market, 193 million bushels of corn changed hands in one day, smashing a trading record that had stood for 122 years. The Dow Jones index for commodity futures hit 145.27, and had its highest one-day advance in nineteen years….
The SCLB epidemic is a dramatic example of an interaction between a crop, whose ‘evolution’ is driven by human intervention, and a pathogen, which evolves naturally to exploit new genetic susceptibilities. The Tcms gene was introduced into maize, by breeders, fewer than 20 years before the epidemic outbreak. Race T likely appeared in the Philippines in 1963, but was not recognized as a new race until the late 1960’s, when it caused the SCLB epidemic in 1970. It is currently unclear whether C. heterostrophus race T evolved around the time of the epidemic or had been ‘lurking’ in the field for a long time. Regardless of the answer, the vast monoculture of Tcms maize was the perfect host for this previously unknown race.
The SCLB epidemic involved the appearance of new race of C. heterostrophus that biosynthesizes a previously undescribed phytotoxin (T-toxin), which renders the fungus highly virulent on T-cms corn. T-toxin is toxic only to the same variety of plant that the fungus is able to attack i.e., maize containing the Texas cytoplasmic male sterility gene (T-urf13). Host-selective toxins like T-toxin are known from about 30 different plant pathogens.
Although much progress has been made towards understanding the genetic nature of the Tox1 locus of C. heterostrophus, the evolutionary origin remains uncertain thirty five years after the emergence of race T. With respect to structure of the locus, the facts are: 1) The Tox1 locus is about 1.2 Mb of DNA distributed on two chromosomes reciprocally translocated with respect to their counterparts in race O. 2) T-toxin production requires at least ten genes (including two polyketide synthases), all of which map to the defined Tox1 locus. 3) Known genes are scattered and embedded in highly A+T rich DNA. 4) Known genes have characteristics typical of other genes involved in secondary metabolism in fungi- they are discontinuously distributed, fast-evolving. Phylogenetic analyses, in general, suggest a history of vertical transmission. There is no compelling evidence for horizontal transfer of the Tox1 genes. 5) Genes mapping to the Tox1 locusare not found in race O. Of 25 PKS genes in C. heterostrophus, only PKS1 and PKS2 are unique to race T. 6) In the genome assembly of race T DNA, known Tox1 genes map to the smallest scaffolds, highlighting the fact that the region is difficult to sequence, due to the A+T rich, repeated nature of the regions surrounding known genes.
Thanks to our ability to do comparative genomics, T-toxin like genes have been discovered recently in few additional fungi as a single compact gene cluster. Recent work has used these Tox1-like genes to build phylogenetic trees to address the issue of evolutionary origin and to ask if any of these additional fungi can produce T-toxin or T-toxin like molecules.
2. Oxidative stress, iron acquisition, and virulence in the Cochliobolus-maize and Setosphaeria-maize interactions.
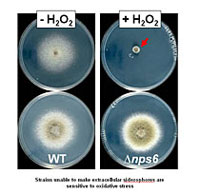 Secondary metabolites and reactive oxygen species (ROS) both have important roles in the dialog between fungal pathogens and their plant hosts. C. heterostrophus is an excellent experimental model to tackle mechanisms of microbe-host communication. The genome carries 12 genes (NPS) encoding non-ribosomal peptide synthetases (NRPS), and one pseudogene. Of the 12 NPSs, only one, NPS6, when deleted, has a virulence function. Strains deleted for NPS6 are also hypersensitive to oxidative and to low iron stress. NPS6 encodes an NRPS responsible for biosynthesis of the extracellular siderophore, coprogen, while a second gene, NPS2, encodes an NRPS responsible for biosynthesis of the intracellular siderophore, ferricrocin (see Project 2). Siderophores are high-affinity iron (Fe+3) chelators; thus, their obvious function is to scavenge and store iron nutrient. They may, however, also act as antioxidants, chelating free iron that would catalyze production of highly cytotoxic ROS. Fungal cells employ conserved signal transduction pathways (STP) to relay information from the environment, including stress, to control gene expression. We have found that mutants in some stress STP genes have phenotypes similar to nps6 extracellular siderophore mutants. Our goals in this project are to examine the interface where small molecule peptide and ROS mechanisms overlap and to determine whether the molecular explanation for common function, is common STPs. Do these pathways, built around sensor kinases, response regulators, and transcription factors link sequestering of iron, production of antioxidants, resistance to oxidative stress, and virulence? The balance between iron as a nutrient and as an agent of oxidative stress may be critical for the outcome of the fungus-plant interaction. ROS are often damaging, yet are now known to provide developmental cues in fungi and thus serve a valuable function. The balance between ROS as a signal and as a stress thus provides another point where virulence could be regulated. The plant and pathogen both produce and detoxify ROS, and they also compete with each other to take up available iron. To unravel this balance we are using fungal mutants in iron metabolism and in signal transduction. We are also asking whether the iron status of the host plant affects infection by wild type and mutant pathogen strains. When we understand the molecular basis for oxidative stress sensitivity and loss of virulence in our siderophore biosynthesis mutants, we may be able to design strategies to manipulate the competition for iron between plant and pathogen to the advantage of the plant. Possible strategies include application of excess iron, application of inhibitors of NRPS activity, or engineering of plants to sequester iron.
Secondary metabolites and reactive oxygen species (ROS) both have important roles in the dialog between fungal pathogens and their plant hosts. C. heterostrophus is an excellent experimental model to tackle mechanisms of microbe-host communication. The genome carries 12 genes (NPS) encoding non-ribosomal peptide synthetases (NRPS), and one pseudogene. Of the 12 NPSs, only one, NPS6, when deleted, has a virulence function. Strains deleted for NPS6 are also hypersensitive to oxidative and to low iron stress. NPS6 encodes an NRPS responsible for biosynthesis of the extracellular siderophore, coprogen, while a second gene, NPS2, encodes an NRPS responsible for biosynthesis of the intracellular siderophore, ferricrocin (see Project 2). Siderophores are high-affinity iron (Fe+3) chelators; thus, their obvious function is to scavenge and store iron nutrient. They may, however, also act as antioxidants, chelating free iron that would catalyze production of highly cytotoxic ROS. Fungal cells employ conserved signal transduction pathways (STP) to relay information from the environment, including stress, to control gene expression. We have found that mutants in some stress STP genes have phenotypes similar to nps6 extracellular siderophore mutants. Our goals in this project are to examine the interface where small molecule peptide and ROS mechanisms overlap and to determine whether the molecular explanation for common function, is common STPs. Do these pathways, built around sensor kinases, response regulators, and transcription factors link sequestering of iron, production of antioxidants, resistance to oxidative stress, and virulence? The balance between iron as a nutrient and as an agent of oxidative stress may be critical for the outcome of the fungus-plant interaction. ROS are often damaging, yet are now known to provide developmental cues in fungi and thus serve a valuable function. The balance between ROS as a signal and as a stress thus provides another point where virulence could be regulated. The plant and pathogen both produce and detoxify ROS, and they also compete with each other to take up available iron. To unravel this balance we are using fungal mutants in iron metabolism and in signal transduction. We are also asking whether the iron status of the host plant affects infection by wild type and mutant pathogen strains. When we understand the molecular basis for oxidative stress sensitivity and loss of virulence in our siderophore biosynthesis mutants, we may be able to design strategies to manipulate the competition for iron between plant and pathogen to the advantage of the plant. Possible strategies include application of excess iron, application of inhibitors of NRPS activity, or engineering of plants to sequester iron.
3. Mechanisms of sexual reproduction in self-incompatible and self-compatible ascomycetes.
 Cellular specialization leading to sexual reproduction is controlled in fungi by mating type (MAT) genes. There are two basic ‘fruitful’ fungal sexual reproductive styles: the first requires partners of opposite mating type whereas the second is accomplished by a single individual without participation of a partner. Fungi that require a genetically distinct partner to complete the sexual process are called ‘self-incompatible’ (heterothallic); species able to undergo sexual reproduction without a partner are ‘self-compatible’ (homothallic). In addition, non-canonical mating strategies, which do not follow the genetic ‘rules’ associated with the best-known mating behaviors have been reported. Our laboratory addresses the structural, functional, and evolutionary issues that correspond to each mating lifestyle. For this, we focus on the species rich Dothideomycete genus, Cochliobolus, as members of this genus are not only diverse in reproductive lifestyle, but also highly amenable to genetic manipulation. Studies tracking the evolution of reproductive lifestyle indicate that homothallic strains arise from heterothallic ones, but direction of ‘evolution’ can be reversed by exchange of MAT determinants.
Cellular specialization leading to sexual reproduction is controlled in fungi by mating type (MAT) genes. There are two basic ‘fruitful’ fungal sexual reproductive styles: the first requires partners of opposite mating type whereas the second is accomplished by a single individual without participation of a partner. Fungi that require a genetically distinct partner to complete the sexual process are called ‘self-incompatible’ (heterothallic); species able to undergo sexual reproduction without a partner are ‘self-compatible’ (homothallic). In addition, non-canonical mating strategies, which do not follow the genetic ‘rules’ associated with the best-known mating behaviors have been reported. Our laboratory addresses the structural, functional, and evolutionary issues that correspond to each mating lifestyle. For this, we focus on the species rich Dothideomycete genus, Cochliobolus, as members of this genus are not only diverse in reproductive lifestyle, but also highly amenable to genetic manipulation. Studies tracking the evolution of reproductive lifestyle indicate that homothallic strains arise from heterothallic ones, but direction of ‘evolution’ can be reversed by exchange of MAT determinants.
4. Role of extracellular DNases in virulence of fungi to hosts. An extracellular DNase from the phytopathogen Cochliobolus heterostrophus is a virulence factor.
This is a collaborative project with Hans VanEtten and Martha Hawes Univ. of Arizona.
During the past decade, new insight into fundamental mechanisms of mammalian immunity emerged when neutrophils were found to function by a process distinct from those believed to predominate in defense (Science 303:1532). Histone-linked extracellular DNA (exDNA) is an integral component of neutrophil extracellular traps (NETs), and when this exDNA is degraded by addition of DNase I, the capacity to immobilize and kill invading pathogens is lost. As a counter defense, bacterial pathogens have been shown to produce extracellular DNases (exDNases): mutation of Group A Streptococcus exDNases results in reduced trapping and systemic spread in animal hosts (Curr Biol 16: 396). Extracellular trapping has now been documented for diverse animal tissues, and shown to play a role in disease response to bacteria, fungi, viruses, and protozoan parasites. We have investigated whether or not a similar mechanism functions in higher plant cell defense. Initially, the Hawes lab demonstrated that plant cells carry out an extracellular trapping process that appears to function in a manner parallel to that of mammalian cells. Histone-linked exDNA can be found as an integral component of a matrix secreted from root caps along with specialized root 'border cells' that trap soilborne bacteria, fungi, and heavy metals. Furthermore, exDNA is required for root tip resistance to fungal infection (Plant Sci 180:741). Our lab has shown that targeted deletion of a gene encoding a putative exDNase in a Cochliobolus heterostrophus is correlated with reduced virulence of the mutant on its plant host. Thus, a fungal exDNase is a virulence determinant. Overall these data provide support for a common defense/counter defense mechanism used by animals, plants and their pathogens.